|
 Download/Print [PDF] Download/Print [PDF]
Concept-Outline |
A Biological Approach to Explaining Morality
|

[ 01 ] |
Scientists are interested in explaining behavior, such as the type exhibited here: why do animals play? Can you name other behaviors that might be addressed scientifically? [Students generate list.] ...Can we include moral behavior on our list? OR What does it mean to include moral behavior on this list?
Target: Orienting to "A Biological Approach to Explaining Morality" |

[ 02 ] |
Here is a pair of blue-footed boobies from the Galapagos Islands: displaying their vivid blue feet in courtship. We can observe the behavior and interpret it in terms of reproduction. We can compare it with courtship behavior in related species. What do we do with morality? Abstract concepts of 'right' and 'wrong', or virtuous motives and good intentions, or systems of justice, do not seem directly observable or measurable. How might you begin to characterize morality as a form of behavior? Can you provide an example?
Target Concept: Morality is a form of behavior. |

[ 03 ] |
Primatologist Frans de Waal notes that "stumptail monkeys (Macaca arctoides) are among the most conciliatory members of the genus Macaca. They are heavily built, yet remarkably friendly and tolerant, such as here: the alpha male is eating attractive food unperturbed by an entire audience around him. When stumptail monkeys were housed with a less tolerant macaque, they modified the latter species' behavior into a more pacific direction."
In what ways might study of macaque behavior inform an understanding of human behavior? What other species you might recommend studying, and why?
Target Concept: Non-human species may exhibit various stages in the evolution of morality. |

[ 04 ] |
How do we define scientifically which behaviors are moral? For example, here a reed warlber feeds a cuckoo chick that has parasitized its nest. Was it "wrong" for the cuckoo's parent to lay its egg in the reed warbler's nest? How do we characterize whether this is a moral question or not?
Perhaps you can suggest a scientific definition of moral behavior? How do we ensure that any such definition does not express a value judgment or introduce bias?
[Follow-up:] Who studies and conventionally defines morality (in a professional sense)? How might one borrow or make use of their expertise?
Target Concept: Biologists borrow from other disciplines in characterizing behavior as moral. |
[ 05 ] |
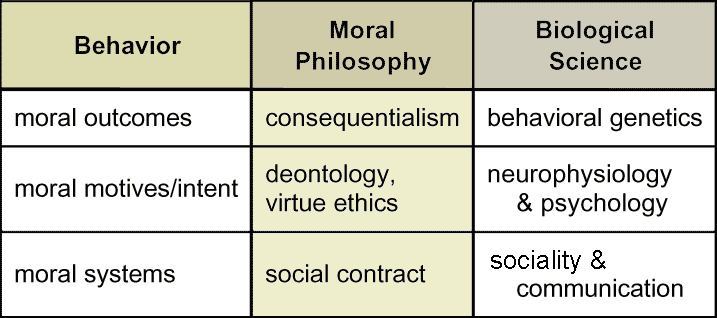
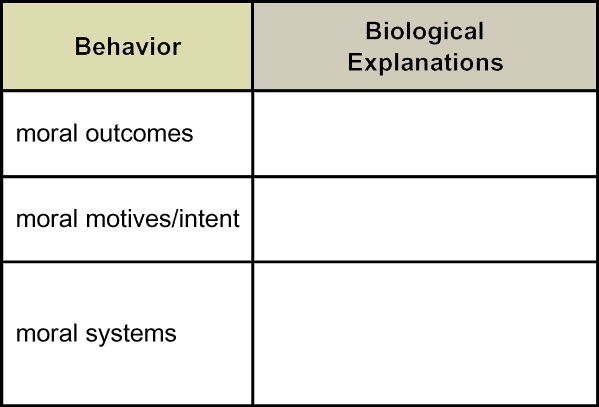
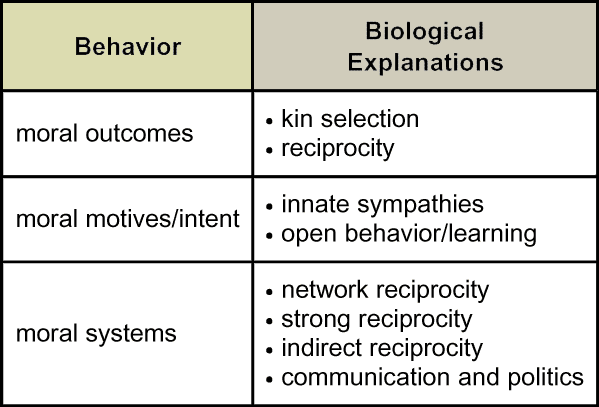
Suppose we turn to moral philosophers. Even after centuries of reflection and debate, they do not all agree among themselves on core ethical principles for defining "good." Generally, however, they recognize three basic approaches. (Does anyone know them?) One approach, consequentialism, focuses on the outcomes themselves. For example, morality is assessed as the greatest good for the greatest number. Good may be defined variously as benefit, happiness or pleasure. A contrasting approach, deontology, emphasizes instead motives or reasoning. For example, morality is measured by feelings of sympathy or virtuous intent. Both approaches draw on widely shared intuitions, although they sometimes lead to different moral codes. They have not yet been synthesized. A third, complementary strand of philosophical thought situates morality on yet another level: the concept of a social contract. Morality is characterized primarily by mutual consensus on values.
Each of these characterizations, or definitions, leads biologists to a different type of explanation. Listed here, then, are three complementary ways one might address questions about the biology of morality. No one explanation may be exhaustive or complete, so all three seem informative.
Any questions or comments before we explore each of these?
Target Concept: Philosophers identify at least three frameworks for conceptualizing morality: moral outcomes, moral motives (or intent), and moral systems. |

[ 06 ] |
Here a langur mother holds a dead baby. Males sometimes kill unrelated infants, increasing the prospects for their own offspring. Natural selection leads to traits that enhance survival and reproduction, so can we say that these values are inherent in nature? Should we consider the male langur's behavior moral (or justified), by virtue of the evolutionary history that led to it?
[Example:] Does a falling body reflect a "value" of gravity?
[Example:] Do two charged particles reflect a "value" of electrical attraction?
=> Does causation make any effect or outcome valued or intended?
[Example:] Some species go extinct. Can we declare survival or perpetuation of the species as "necessary" or an inherent value of evolution?
[Example:] Some insect castes are sterile. Other individuals are infertile. Can we declare reproduction as "necessary" or an inherent value of evolution?
=> What is the relationship between the outcome of evolution and intepreting intention or purpose in the process?
Target Concept: Evolution itself does not express or yield values. (Evolution is non-teleological.) |

[ 07 ] |
If biologists can explain moral behavior, can they then justify certain moral principles? In studying morality biologically, can we hope to ultimately develop a truly scientific system of ethics based on our observations? Why or why not?
Suppose we find some emotional disposition among humans to help (or to harm) others: in what ways would this help guide our ideals or judgments of moral behavior? Does it matter if the disposition is found to be universal? What if there are exceptions?: Can you have "unnatural" products of nature?
How should we characterize the difference between description and prescription? What are the aims and scope of scientific analysis? Do we need to endorse specific moral principles to understand why or how they are exhibited?
Target Concept: Science is limited to description. |
Moral Outcomes (Behavioral Genetics)
|

[ 08 ] |
The first category of biological explanations for moral behavior (listed in Table 1) involves interpreting the acts or outcomes in an evolutionary context, in terms of fitness, or survival and reproduction measured in genetic terms. Some behaviors or dispositions — not all — are partly hereditary (innate or instinct). To the degree that they are, they will be subject to natural selection. Based on your understanding of natural selection, what kind of behaviors will result? Does cooperation or helping others seem possible?
Consider this example: insect pollination of flowers. The flower produces nectar at a cost, but the insect benefits: why?
Target Concept: Cooperation and helping behavior are simple evolutionary puzzles. |


[ 09 ] |
Here is an example of cooperation, or mutualism, between species: the ant/acacia relationship. The tree has large thorns, which serve as nests; nectaries, which provide sugars for the ants; and Beltian bodies, which provide protein. The ants patrol the tree and the ground underneath it, eating plants that might take light or water from the acacia, removing fungal spores, and stinging any potential herbivore. How might this system have evolved, in steps, from ancestors that had no such cooperative relationship? Why has the cooperation developed, even with increasing "costs" for each organism?
Can you name any other examples of mutualisms or systems of mutual benefit or exchange?
Target Concept: Organisms may cooperate when each benefits. |

[ 10 ] |
Mutualisms exist between species. What about between organisms within the same species? Such organisms will tend to use (and compete for) the same resources. Under what circumstances might they cooperate, as is the case of these musk oxen, in a defensive formation?
[This case was about prey. Can you identify circumstances in which predators, too, might benefit from cooperation?]
Target Concept [continued]: Organisms may cooperate when each benefits. |

[ 11 ] |
Here is one more example of intraspecific cooperation. When cliff swallows go out again after returning from a successful foraging event, other birds sometimes follow them. Information about the location of food is thus shared. If the birds were competing for the same food resources, what type of behavior would you expect to be most adaptive? In this case — also observed among osprey, weaver birds, crows, honeybees, ants, termites and others — food becomes available in periodic, plentiful batches. Food is not limited by competition, and the information about where to find it may be shared at no relative cost to the individual. Individuals also benefit when they can freely follow others.
Target Concept [continued]: Organisms may cooperate when each benefits. |

[ 12 ] |
Here's an interesting nest: The chick is 18 days old. The father perches above. But on the nest are both the mother and the chick's older sibling. The sibling is old enough to reproduce herself: why is she helping to raise her sibling, rather than having a nest of her own? (Can you propose any hypotheses?)
When Florida scrub jays (Aphelocoma coerulescens) reach reproductive age, they do not always leave home and begin their own families. A male scrub jay must have his own territory for foraging and nesting, and territory is limited. Imagine a male who stays with his father. He can help gradually expand the father's territory. How might this benefit the son?" [Eventually, the territory can be split between father and son. This way the son is better able to secure good territory.]
"Females, by contrast, compete for males with the best territories. How might staying at home be beneficial?" [A female who can wait for opportunity is able to select a better mate.]
In each case, the scrub jay actually benefits reproductively in the long-term by staying at home and helping their parents reproduce in the short-term (Woolfenden and Fitzpatrick 1978, 1984). When viewed in a larger context, an apparent cost actually increases their own reproductive potential, and is favored by natural selection.
Reproductive helpers are also found in black-backed jackals (Canis mesomelas), cichlid fish of Lake Tanganyika (Lamprologus brichardi), grey-crowned babblers of Australia (Pomatostomus temporalis), carrion crows in northern Spain (Corvis corone corone) and many other species (Krebs and Davies 1993, pp.299-302; Clutton-Brock 2002; Baglione et al 2003).
Target Concept: Some cases of "costly" helping are apparent only. |

[ 13 ] |
Here is another case of helping: sentinel behavior in meerkats (Suricata suricatta) (Clutton-Brock et al 1999). Meerkats are a type of mongoose found in arid southern Africa. They typically live and forage in groups. Some individuals watch for predators and sound a general alarm if one is spotted. By vocalizing loudly, however, the sentinel seems to alert any nearby predator and put herself at more risk: an apparent costly act. Can you imagine any circumstances where this might be beneficial? . . .
Sustained observation reveals that meerkats guard from safe vantage points, where they can readily escape into a burrow. In addition, sentinels are usually the first to detect the predator and they hide sooner than most others. Vigilance limits foraging time, however. Meerkats tend to adopt a guard role only when they are well fed.
The same pattern of sentinel volunteering has been observed in the bird, the Arabian babbler (Turdoides squamiceps) (Wright et al 2001), and others.
Target Concept [continued]: Some cases of "costly" helping are apparent only. |

[ 14 ] |
Let's consider how a larger context might inform analysis of other cases of costly helping, as well. Honeybees, and many wasps and ants, along with naked mole rats from eastern Africa include individuals that do not reproduce. Rather, they contribute to the reproduction of a single individual in a social setting. The failure to perpetuate one's own lineage seems to contradict the principle of natural selection. But consider the more inclusive lineage of collateral relatives. Contributions to the survival and reproduction of kin may well outweigh the individual's own. Costly helping traits might be preserved and proliferate through relatives, not direct offspring. Natural selection would be indirect: kin selection. The benefits would depend, of course, on the degree of relatedness, or the likelihood that the helping genes were passed on through relatives. How much genetic information does an individual share with its offspring? —with its siblings? —with its siblings' offspring? —with its cousins? Using this information, what combination of benefits to others would (genetically) outweigh one's own cost?
The genetics of honeybees, pictured here, are unlike humans'. Females are diploid, while males are haploid (developing from unfertilized eggs). Accordingly, a female is generally 75% related to her sisters, 50% to her own offspring and brothers. How would you expect this to affect the natural selection of helping and reproductive behavior in females?
[Honeybee hives are indeed composed of groups of cooperating sisters, or workers, all descended from a common mother, the "queen bee". Males are not part of the hive. The female workers are sterile.]
Target Concept: Some cases of costly helping are explained by genetic relatedness. |

[ 15 ] |
Naked mole rats (Heterocephalus glaber) live in underground colonies, an adaptation to patchy food resources in an arid environment (O'Riain and Faulkes 2008). Some colonies are relatively isolated and, due to inbreeding, individuals in the same burrow typically share at least 80% of their genes. How might this affect the natural selection of reproduction and helping behavior?
[In naked mole rate colonies, only one female, much larger, is reproductive. Other females become sterile (Jarvis 1981; Sherman, Jarvis and Alexander 1991).]
Target Concept [continued]: Some cases of costly helping are explained by genetic relatedness. |

[ 16 ] |
Belding's ground squirrels (Spermophilus beldingi) inhabit mountain meadows in the western U.S. Helping behavior appears in the alarm calls of sentinels watching for predators, an evolutionary result of foraging in the open during the day. Unlike meerkats, however, Belding's ground squirrels that raise an alarm are more frequently preyed upon. Their social system also differs. Males tend to disperse from their place of birth, while females remain in the local area. How might this affect the selection of sentinel behavior?
[Females have more kin neighbors than do males and, accordingly, they devote more time to the sentinel role. In addition, their alarm calls are more numerous when only close relatives are nearby (Sherman 1977; Shelley and Blumstein 2004).]
Target Concept [continued]: Some cases of costly helping are explained by genetic relatedness. |

[ 17 ] |
Kin selection can only be effective, of course, if organisms can differentiate kin from non-kin (Grafen 1990). Since they cannot determine each other's genes directly, kinship or degree of genetic relatedness must be perceived indirectly, through vicarious clues (Campbell 1974). In what ways might organisms be able to identify or detect kin, directly or indirectly? [Students list possibilities.]
Here are a few examples. In the case of Belding's ground squirrels, locality itself may function as a substitute when kin tend to inhabit the same area. Boobies on the Galápagos Islands (Sula nebouxii), pictured here, also recognize their offspring only by location: within a ring of guano that marks their rudimentary nest. If a nestling falls outside the ring, whether accidentally or by being pushed, it is treated as non-kin and is neither cared for nor permitted to return (Gould 1983).
Target Concept: Organisms can determine kin relatedness in many ways, sometimes by indirect cues. |

[ 18 ] |
American cowbirds (here) and European cuckoos lay their eggs in the nests of other species, where the host birds raise the chicks as if kin. What makes this nest parasitism possible?
Target Concept [continued]: Organisms can determine kin relatedness in many ways, sometimes by indirect cues. |

[ 19 ] |
Side-blotched lizards in California (Uta stansburgiana) have three throat colors: orange, yellow and blue. They establish mutually favorable territories based on the throat visual cues, even when unrelated individuals are mixed. Here, the trait seems to signal genetic similarity, even if the individuals are not fully related genetically (Sinervo and Clobert 2003).
The ability of humans to ascertain kin has been investigated recently with special regard to attitudes towards incest and its avoidance. Primates, including humans, can track kin relationships through observed births — of younger (but not older) siblings. They use this information in social interactions (even if unconsciously). In other cases, siblings seem to use years of cohabitation, regardless of whether they are actually kin, as a vicarious indicator (Lieberman, Tooby and Cosmides 2003, 2007). While humans in most cultures tend to associate in family groups, the role or extent of kin selection remains an open question.
Target Concept [continued]: Organisms can determine kin relatedness in many ways, sometimes by indirect cues. |

[ 20 ] |
Vampire bats (Desmodus rotundus) cannot survive without food more than a few days. But on any given night, 7-30% of the bats fail to find a meal. A bat may then turn to a roostmate and nuzzle its throat. On roughly 5 of 8 occasions, the second bat regurgitates a small amount of blood for the first bat. This occurs even among non-kin (Wilkinson 1984, 1988, 1990). Can you imagine a context in which this costly helping behavior might evolve?
[Note: bats frequently return to the same roost — and roostmates.]
[Vampire bat sharing behavior is not an isolated act. Eventually, the donor bat fails to find food. Roles of beggar and potential donor are then reversed. Here, exchanges (as in other cooperative mutualisms) may simply be displaced in time. Namely, the recipient may later reciprocate, essentially closing an unstated deal (Trivers 1971).]
Grooming is not critical to survival, but it occurs frequently in many primate species, even among unrelated individuals. It is typically reciprocated by later grooming or other social "favors." In one study of food sharing in a chimpanzee troop, based on over 7,000 observed interactions, exchange was ultimately balanced for each chimp pair (de Waal 1989).
Target Concept: Some cases of costly helping are explained by reciprocity. |

[ 21 ] |
Among black hamlet fish of the Carribean (Hypoplectrus nigricans), the potential exchange involves reproduction. The fish are hermaphroditic. They can both spawn and fertilize. The energy investment in eggs is much greater than in sperm, however, so the reproductive cost to the fish that lays eggs is greater. What strategy would you recommend to a black hamlet to ensure reproductive reciprocity with an unknown individual encountered on the reef? That is, how might you guard against a cheater, and ensure that the other individual fulfills its end of the "bargain"?
[For black hamlets, any initial laying of eggs is limited. Successive matings depend on the alternation of male and female roles (Krebs and Davies 1993, p. 285). Here, as elsewhere, repeated encounters allow reciprocity.]
Arrangements of reciprocity are indeed fragile and susceptible to cheaters. Such relationships have been modeled using computers for repeated encounters among individuals using various postures of cooperating or "defecting." An individual that "reciprocates" by doing whatever the other individual has done (whether to cooperate or not) seems to be the most effective strategy (Axelrod 1984).
Target Concept [continued]: Some cases of costly helping are explained by reciprocity. |
Moral Motive/Intent (Neurophysiology & Psychology)
|

[ 22 ] |
This is a macaque, or rhesus monkey. Who is wondering what this animal might be thinking? How does this differ from thinking about the macaque's behavior genetically? What is important when addressing morality: the outcomes alone, or also the motive or intent? (Recall the various philosophical perspectives in Table 1.) How would a scientist go about trying to study such intangible internal states of mind?
Target: Orienting to neurophysiology and psychology as a way to address motive and/or intent as dimensions of moral behavior. |

[ 23 ] |
This 1862 painting by Adolphe Bourguereau portrays a scene from Greek mythology, "The Remorse of Orestes, or Orestes Pursued by the Furies." Orestes was persuaded by his sister, Electra, to kill his mother to avenge the murder of their father, which he does in a fit of passion -- only to later regret his action. The sentiment of remorse was considered by Darwin and others of his period to be deeply expressive of moral sentiments (Descent of Man, p. 91.) What elements might be required historically for an organism, such as humans, to be able to experience such remorse or conscience? How, from a biological or evolutionary perspective, might we have developed the capacity to feel moral duty?
Charles Darwin proposed four interrelated features. First, Darwin observed that animals could evolve societies, structured (he assumed) by a social instinct. Second, with multiple instincts, behavior might not always accord with social benefit. But memory, Darwin thought, would help resolve such conflicts as the organism learned to regulate its instincts, making the social instinct primary. Third, the use of language would allow organisms to communicate their needs clearly to one another. Fourth, repetition would lead to habit, and a spontaneous sense of what one "ought" to do.
In what ways do you find this account informative? —In what ways insufficient or incomplete?
Target Concept: Darwin proposed the moral sense as an inevitable outcome of four elements: social instinct, memory, language and habit. |

[ 24 ] |
In 1996, Binti Jua, a female gorilla at the Brookfield Zoo outside Chicago, rescued a three-year-old child that had fallen into her enclosure. Binti Jua cradled the unconscious boy, just as she did her own child, growled at another gorilla who approached her, and then carried the boy about 20 meters to a door where the zookeeper could retrieve the boy, all while her own daughter clung to her back (Bils and Singer 1996; "Gorilla at an Illinois Zoo..."). The incident echoed a similar case at England's Jersey Zoo in 1986 with a male gorilla named Jambo. What do you regard as an appropriate conclusion about the gorilla's intent or motives? Do you regard this as an expression of sympathy in another primate?
Target Concept: Non-human organisms exhibit sympathetic concern for both kin and non-related individuals. |

[ 25 ] |
This is Mozu, a snow monkey, who was born without hands or feet in a free-ranging troop in Japan. Mozu could not climb. Still, the other members of Mozu's troop did not move in the trees above her at a pace that would leave her behind, despite their ability to do so. Why might they have tolerated Mozu's handicap? What might this indicate about human emotions of sympathy?
Target Concept [continued]: Non-human organisms exhibit sympathetic concern for both kin and non-related individuals. |

[ 26 ] |
In an effort to understand the nature of sympathy in non-humans, in 1963 Jules Masserman investigated how macaques, such as these, responded to other monkey's suffering in a laboratory environment. How would they behave if they knew that securing food would give an electric shock to another monkey? (Changes in research ethics no longer allow such studies.) What would you expect, here?
Masserman's monkeys often prolonged their hunger rather than administer a painful stimulus. One monkey refrained from taking food for twelve days. Responses showed several patterns. Self-starvation was more likely in monkeys that had themselves experienced electroshock as a subject. Sacrificial behavior was not biased towards members of higher dominance rank, but was slightly stronger for cagemates (although not statistically significant). Visual contact, even without auditory cues, seemed sufficient to induce the response. What conclusions about sympathy in non-human primates would you draw from this study? For example, did the monkeys' behavior seem to reflect an understanding of and concern for another's pain?
Mice also seem to show signs of proto-empathy in expressing feelings modified by the feelings of others. Experimentally, they exhibit increased sensitivity to mild pain when cagemates (but not unfamiliar mice) also experienced noxious stimuli at the same time. Again, visual contact seems important in communicating an emotional state and triggering a corresponding, even if not directly empathetic, response (Langford et al 2006).
Target Concept [continued]: Non-human organisms exhibit sympathetic concern for both kin and non-related individuals. |

[ 26a ] |
In 2008 Frans de Waal performed a related study on capuchin monkeys, involving food rewards rather than painful stimuli. As pictured here, the capuchins were presented with two options: one selfish and one that provides food to themselves and also to a nearby monkey. What option do you think they chose? —Under what circumstances?
The monkeys chose the prosocial option when the other monkey was visible and familiar, and so long as the other monkey did not receive a larger portion of food (de Waal, Leimgruber & Greenberg, 2008).
Target Concept [continued]: Non-human organisms exhibit sympathetic concern for both kin and non-related individuals. |

[ 27 ] |
Common marmoset monkeys (Callithrix jacchus) are cooperative breeders and are strongly interdependent socially. In one test, they were given an opportunity to provide food to other individuals, but with no personal reward. What do you think they chose to do?
In these cases (even without reciprocity or genetic relatedness) they extended a small effort to provide food for others (Burkart et al 2007). In what ways might these findings about unsolicited, other-regarding behavior in apes, snow monkeys, macaques, capuchins and marmosets inform our understanding of human moral sentiments?
Target Concept [continued]: Non-human organisms exhibit sympathetic concern for both kin and non-related individuals. |

[ 28 ] |
"These rhesus monkeys are only a few month old. The smaller monkey in front, a female, has just been molested by an adult" (de Waal 1996). What might be indicated by the other infant monkey (a male) hugging her?
Such acts of apparent consolation are not observed in older monkeys. Would you consider this behavior innate or learned? What might this indicate about human children and their observed spontaneous expressions of sympathy or help?
Target Concept: Humans and some other primates exhibit moral sentiments at a very early age. |

[ 29 ] |
This chimp is helping the human reach something inside the box by lifting a panel the human cannot reach. This was part of a recent study that showed spontaneous (unsolicited) help in young chimps (ages 3 to 4˝ years), as well as in human infants (age 18 months) [Warnecken & Tomasello 2006]. Would you consider such helping behavior primarily innate or learned? What does it mean that such helping behavior is less common in older chimps?
Target Concept [continued]: Humans and some other primates exhibit moral sentiments at a very early age. |
[ 30 ] |
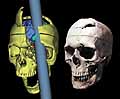
If evolutionary understanding is informed by analysis of structure and function, we might expect to further understand moral behavior by considering the anatomy and physiology of the brain and nervous system (Pfaff 2007). Illustrated here is a fascinating case of an unusual brain injury to a railroad worker in the 19th century, Phineas Gage. Gage was using his tamping iron to compact an explosive charge when it detonated prematurely and sent the 1˝-inch-wide rod up under his cheek bone, through his brain, and out the top of his skull (as reconstructed at left). Gage, remarkably, survived (his partially healed skull, at right). But he had lost some of his brain, and with it, some of its function. Whereas before the accident Gage had been "quiet and respectful," afterwards he became "fitful, irreverent, indulging at times in the grossest profanity (which was not previously his custom), manifesting but little deference for his fellows." The attending physician profiled the dramatic change, noting that "the equilibrium or balance, so to speak, between his intellectual faculties and animal propensities, seems to have been destroyed" (Harlow 1868, pp. 339-340). What should we conclude about the relationship of brain structure and moral function based on this case?
Target Concept: Moral thinking and feeling have a neurological basis. |

[ 31 ] |
In Gage's time, phrenology was a popular theory claiming that discrete personality traits mapped onto the surface of the skull, as viewed in this model. One enthusiast imagined that in Gage's brain "the iron had passed through the regions of the organs of benevolence and veneration . . . hence his profanity, and want of respect and kindness" (Macmillan 2000, quote on p. 350). Today phrenology has long since been abandoned in disrepute, with little evidence for such personality divisions within the brain, nor their correlation with skull shape. What should we conclude about the possibility of specific areas of the brain specializing in exclusively moral mental processes, or the power of science to answer such questions reliably?
Target Concept [continued]: Moral thinking and feeling have a neurological basis. |
[ 32 ] |
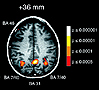
Here is an fMRI showing brain areas used by a subject engaged in thinking about a personal (rather than impersonal) moral problem. Such neurological imaging techniques have proven useful in recent years in monitoring brain activity related to moral thinking and feeling. Such studies seem to show that, in practice, moral thinking seems to involve both emotion and logic (Greene and Haidt 2002). Philosophers have disagreed for centuries about which is (or should be) primary in moral judgment. In what ways might these studies contribute to interpreting that long-standing debate?
Target Concept [continued]: Moral thinking and feeling have a neurological basis. |
[ 33 ] |
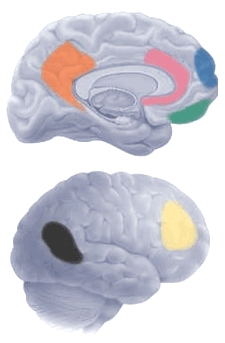
Using the neurological imaging technology, several specific brain areas are found to relate to particular mental functions (or groups of them), although those functions cannot yet be defined precisely. For example, the ventromedial prefontal cortex (green) shows activity when subjects view images that evoke moral impressions but do not require any actual moral judgment. This area has been proposed as part of a network involving feelings related to social interactions. People with damage to this area (such as Phineas Gage) are less able to integrate emotional information into judgments that also involve an analysis of costs and benefits, and their conclusions reflect a corresponding bias. By comparison, when moral reflection turns to interpreting and assessing other people's intentions, the key brain area seems to be the right temporoparietal junction (brown). When moral problems become personal, the medial frontal gyrus (blue) becomes more active. More abstract or hypothetical problems, by contrast, tend to activate the dorsolateral prefontal cortex (yellow) and other areas. When such different forms of thinking conflict, as one often finds in moral dilemmas, activity rises in the anterior cingulate cortex (pink) — perhaps serving a mediator role. The posterior cingulate (orange) seems involved in integrating emotions, imagery and memory, especially import to coherent narratives. All regions active in moral thinking have been implicated in other, non-moral mental processes (Damasio et al 1994; Greene and Haidt 2002; Miller 2008).
Based on these studies, how should one interpret the concept of a "moral organ"? How should we characterize the nature of moral thinking (as compared to other types of thinking)? What implications might these findings have for interpreting moral behavior genetically?
Target Concept [continued]: Moral thinking and feeling have a neurological basis. |

[ 34 ] |
Here, one chimpanzee demonstrates for another how to crack hard panda nuts with a stone hammer on a natural anvil. This is not just tool use, but also teaching and learning. How does this differ from innate behavior? [Clarify closed vs. open behavioral programs.]
What potential benefits does learning [or open behavior programs] bestow, in contrast to innate behaviors? How will this affect the evolution of neural structures that enable learning?
How might this apply to moral behavior? Which aspects of human moral behavior seem primarily learned, which primarily innate? What evidence do we have to make these judgments?
Target Concept: Moral behaviors may be learned as part of an open behavioral program. |

[ 35 ] |
Consider the consequences of learning for evolution. How will selection act on learned behavior? Is it possible for specific learned behaviors to be selected? (In what way?) How is fitness exhibited or "measured" in such cases? How does the ability to learn itself evolve?
In what ways might learning be able to modify innate behavior? How should we charaterize the relationship between behavioral genetics and mental (or psychological) functioning, on two apparently diferent levels of organization?
Imagine two genetically identical animals: will their learned behavior also be identical? What does this tell us about the "identity" of organisms with open behavioral programs?
How would you characterize behaviorally the notion of choice? What makes genuine choice possible? What should a biologist say about the concepts of intention, agency and free will?
Target Concept [continued]: Moral behaviors may be learned as part of an open behavioral program. |

[ 36 ] |
Here, a laboratory setting enables chimpanzees to work together to secure food: they must both pull a rope to bring a board carrying the food within reach. Sometimes they collaborate, sometimes not. Given a choice, the chimps recruit peers with the best history of cooperation (Melis, Hare & Tomasello 2006). Similar cooperative and recruiting behavior has also been documented in the crow-like rooks of Europe (Corvus frugilegus) (Holden 2008; Seed et al 2008). Do you regard this as evidence that the chimps and rooks understand cooperative behavior, and that their actions are intentional? Given that we cannot read another person's — or another organism's — mind, how do we make conclusions about intention?
How might the ability to learn, or intention, be relevant to the problem of defection or cheating in a system of reciprocity?
Target Concept: Cooperation and helping behavior may be intentional. |
Moral Systems (Sociality & Communication)
|

[ 37 ] |
Behavioral genetics alone does not solve the problem of how reciprocity might function in a social setting. Psychological level considerations alone leave open the question of what may be learned from others. So we might turn to the third conception of morality, moral systems (Table 1).
A closed genetic program or psychological predisposition to help or exhibit reciprocity has a potential flaw (which you may have already noticed?): what about cheaters? Won't selfish individuals proliferate by "free-riding" at the expense of others? Won't that subvert any system of cooperation or sharing? Here is one possible example: the ozone hole, typical perhaps of many environmental problems. What prevents someone from using more than their fair share of a shared resources, or "commons," such as public lands, rivers, oceans, the air. Philosopher Garret Hardin (1968) called this problem the tragedy of the commons. Economists encounter this problem when they discuss public goods, intended to be distributed evenly, but freely available for anyone to take a disproportionate share. What are the conditions under which this problem develops? Can you imagine any solutions to this problem? How might the abilities to learn and to interact with others in a social context alter the dynamics of cooperative or selfish behavior?
Target: Orient to moral systems, and the roles of sociality and communication |
[ 38 ] |
Consider again the vampire bats. Their observed system of blood exchange seems stable, undisrupted by potential free-riders. Why?
The bats tend to roost in the same colony, a very simple social organization. How might this affect potential cheaters?
If you were a vampire bat, how would you respond to an individual who was seeking a share of blood for a second or third time? How would you respond to an individual who had not provided you blood after one of your unsuccessful foraging outings? Indeed, the bats do not always provide blood when "begged" from another. They refrain more frequently in just these cases. A bat that does not repay previous "favors" does not get endless handouts (Wilkinson 1984, 1988, 1990). What consequences does this have for cheaters? What conditions or capacities are needed for the bats to be able to maintain their system of reciprocity?
[ The bats illustrate the general principle that organisms may cooperate selectively with reciprocators, aided by memory and the recognition of individuals. Selective interaction leads to a social cluster of cooperators insulated against invasion by selfish individuals: a concept known as network reciprocity (Nowak 2006). ]
Target Concept: Organisms may cooperate selectively with reciprocators. |

[ 39 ] |
A system of sharing is also found in a free-ranging (semi-captive) colony of macaques, or rhesus monkeys (Macaca mulatta), on an island off Puerto Rico. Individuals call to the group when they find food. However, some individuals fail to call. When they are discovered they receive adverse treatment. They are more likely to be bit, hit, chased or rolled (Hauser 1992). What are the consequences for cheaters and cooperation here?
What are the conditions or capacities that enable such a system of punishment or enforcement?
[ Cooperation enforced through punishment is called strong reciprocity. ]
Target Concept: Social organisms may enforce cooperation through rewards and punishment. |

[ 40 ] |
What about strong reciprocity among humans? When Darwin began considering the evolution of morality, he reflected on a possible role for the "fear of others acting in unison" and "the fear of punishment" (M Notebook, p. 151; 1871, p. 92). However, consider that punitive behavior involves a cost of extra effort or resources. Do you think humans accept these added costs?
Recent studies do show that individuals, even in anonymous experimental situations, do accept personal cost to ensure group benefits. Moreover, others respond to their punitive actions (Fehr and Gächter 2002). Punishment of selfish behavior seems present in all human cultures. They include not only different nations on different continents and Oceania, but also cultures with widely divergent environments, economies (from foraging and pastoralism to industrialism) and residence patterns (from nomadic to sedentary) (Heinrich et al 2006; Herrmann, Thoni and Gächter 2008). Most notably, perhaps, negative sanctions are found in small mobile hunter-gatherer cultures — similar to our Paleolithic ancestors (Boehm 1999). What do these findings imply for the origin and maintenance of an egalitarian society, such as is typically found in such hunter-gatherer cultures (but not among chimpanzees)?
Target Concept [continued]: Social organisms may enforce cooperation through rewards and punishment. |

[ 41 ] |
What other ways might organisms regulate the occurrence of cheating? Here is the small cleaner fish Labroides dimidiatus, which has a mutualism with large fish. The small fish eats parasites on the large fish. Occasionally they "cheat" and feed on the host's mucus or nip a bit of its flesh. Other potential host fish (or "clients"), however, can observe or "eavesdrop" on such behavior. How might this affect the dynamics of cheating and "honest" reciprocity?
In fact, host fish show a preference for cleaner fish that are demonstrably "honest." Accordingly, cleaner fish cheat less when bystanders are present (Bshary and Grutter 2006). The use of social information to evaluate other organisms' behavior patterns is known as image scoring.
Reputation can matter, it seems. Cooperation can be guided by status, or social information, rather than instances of direct reciprocity. Indirect reciprocity can evolve in a group with image scoring (Nowak and Sigmund 1998, 2005) — and can also effectively solve the problem of the tragedy of the commons (Milinski et al 2002).
Target Concept: Organisms may benefit from social information. |

[ 42 ] |
Given the examples already noted, how might you characterize the role of cognitive and social contexts in shaping cooperative behavior?
Most monkeys do not help others unless there is a personal benefit. That might be due to their limited cognitive ability. However, common marmosets (Callithrix jacchus) retrieve food for others in experimental situations, even with no clear benefit to themselves. Unlike closely related species, the marmosets breed cooperatively (Burkart et al 2007), a social system shared with the great apes. [continued...]
Target Concept: Variations in social and cognitive contexts shape helping and cooperative behavior. |

[ 43 ] |
[...continued]
Both chimps (primates) and rooks (birds) can recruit helpers to perform joint tasks. But their behaviors also differ, as do their social organizations. Rooks do not seek or achieve cooperation as frequently as chimps. Their social organization is also simpler. They form single mating pairs that tend to remain for life. Chimps, by contrast, shift mating partners and must also negotiate political allegiances in an unstable dominance hierarchy. In addition, cooperation among individual pairs of rooks reflects their mutual tolerance in other social encounters.
In humans, punishment occurs more readily in societies with stronger norms of cooperation (Heinrich et al 2006; Herrmann, Thoni and Gächter 2008).
Given all this information (about marmosets, chimps, rooks and humans), how would you characterize the respective roles of cognitive or social context in shaping moral behavior?
Target Concept [continued]: Variations in social and cognitive contexts shape helping and cooperative behavior. |

[ 44 ] |
There are also significant differences in social structure between early human ancestors and their closest primate relatives. Here, "three male chimps [Pan troglodytes schweinfurthii] share the carcass of a red colobus [Colobus tephrosceles badius] which they have caught and killed. A female with ventral infant (right) begs for scraps of meat" (Bygott). The scene reflects how sharing in chimps is both motivated by and limited by political context. Chimps compete for both food and mates, even within social groupings. Their societies are marked by linear dominance hierarchies. Pairs sometimes form coalitions and significantly alter the balance of power. Larger coalitions appear temporarily but they are limited in scope. Chimp cooperative behavior is limited, and often politically oriented. Altruism is rare. [continued...]
Target Concept [continued]: Variations in social and cognitive contexts shape helping and cooperative behavior. |

[ 45 ] |
[...continued]
Early humans (around 100,000 years ago) seemed able to level dominance hierarchies (still found today among chimpanzees). Communication skills facilitated the coordination of large coalitions that could effectively check the authority of dominant individuals. Weapons, once developed for hunting, likely contributed further to equalizing power. Egalitarianism emerged and, with it, moral norms (Boehm 1999). What factors (cognitive, social, technological, other) seem most significant in shaping the evolution of early human morality? What might have initiated those changes?
[Advanced:] In what ways does the evolution of human morality seem "inevitable," in what ways contingent on certain historical events?
Target Concept [continued]: Variations in social and cognitive contexts shape helping and cooperative behavior. |


[ 46 ] |
In Descent of Man, Darwin identified the development of language as one of the four elements central to the development of human moral sentiments. (The others were social instinct, memory, and habit.) In what ways might communication be essential to or contribute to moral behavior, or (for Darwin) to responses based on moral sentiments or feelings?
Darwin followed Descent of Man with a whole volume exploring non-linguistic communication: The Expression of Emotions in Humans and Other Animals (1872). Here are some samples: a "hostile" dog and a "humble and affectionate" dog illustrate his principle of antithesis -- that opposite emotions are associated with opposite postures or muscles. . . . [continued...]
Target Concept: Effective communication enhances the social function of moral behavior. |
[ 47 ] |
[...continued]
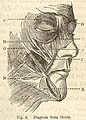
. . . Darwin also analyzed the muscular anatomy of facial expressions, . . .
[continued...]
Target Concept [continued]: Effective communication enhances the social function of moral behavior. |
[ 48 ] |
[...continued]
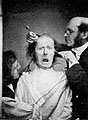
. . . in part, drawing on the experiments of G.B. Duchenne, who used electrodes to stimulate particular muscles and then photographed the resulting facial expressions, linking them to the emotions they conveyed. . . .
[continued...]
Target Concept [continued]: Effective communication enhances the social function of moral behavior. |

[ 49 ] |
[...continued]
. . . Contemporary studies continue to document the role of particular muscles in expressing emotions, as well as in feeling them and perceiving them among others (Niedenthal 2007). In what way might understanding the expression of emotions contribute to understanding moral behavior in a social context? What else might need to be communicated, or discerned, besides emotions?
Based on these images about the communication of emotion, how might organisms interpret the intention of others, so important to many moral judgments? How might one organism deceive another about its intentions, and how might that other organism detect such deceit? [These problems are still being investigated.]
What other aspects of communication might shape the sociality of moral behavior?
Target Concept [continued]: Effective communication enhances the social function of moral behavior. |

[ 50 ] |
Society is often viewed as the simple sum of individual interactions. But consider how social interactions may, in turn, shape individual behavior and even biological fitness. Here is one example: the awarding of a medal to a journalist wounded in wartime-reporting. The award conveys publically how the community values service and sacrifice. Such social rewards, like punishment too, help shape what an indvidual learns. Can you identify other possible examples where social context may foster, discourage or shape moral behavior? Some possibilities to explore: expressions of sympathy? readiness to punish? tendency to help? language ability? perceptual abilities? depth or acuity of memory? What effect might these various contexts have had earlier in human evolutionary history? How might these factors vary in family groups, small multi-family groups or larger tribes?
Target Concept: Social interactions may affect individual learning and biological fitness. |
Summary & Application
|
[ 51 / 51a ]
|
Having now considered biological perspectives on morality on mutiple levels — behavioral genetics; neurophysiology and psychology; and sociality and communication — how might we summarize how biologists explain the philosophers' dimensions of moral behavior: moral outcomes, moral motives or intent, and moral systems? What concepts are central to each of these explanations? How do these three sets of explanations relate to each other?
Target Concept: Biologists can explain morality on multiple levels. |

[ 52 ] |
In popular culture, biology – including behavior — is typically reduced to genes, as exemplified in this Time magazine cover headline. What examples in this presentation limit that conception for moral behavior? For example, what are the roles of learned behaviors at the psychological level and of regulation of behavior through interactions at the social level?
In what ways do such perspectives of biological determinism shape assumptions about the biology of morality? How might any such misleading assumptions in our culture be remedied?
Target Concept: Higher levels of organization limit reductionistic explanations of behavior. |

[ 53 ] |
Natural selection is often described using the phrases "struggle for existence" and "survival of the fittest." What meanings and connotations are conveyed in these phrases? Can you identify other ways our culture portrays selfishness or ruthless competition as "natural" or inevitable? How might the widespread use of these images (and other expressions, such as "law of the jungle" or "dog-eat-dog world") affect popular perceptions about nature and about the propsect that human morality developed through evolution? How do these popular perceptions compare to current biological understanding of morality (on all three levels)?
For example, what is the role of: mutualisms between species, reciprocities among individuals within a species, and the potentials of open behavioral programs, innate sympathies, social networks of reciprocity, punishments and rewards, image scoring (or reputation), and social contexts?
Target Concept: Cultural images of nature, natural selection and evolution tend to bias how human morality is viewed in a naturalistic perspective. |

[ 54 ] |
In the late 1800s self-styled philosopher Herbert Spencer claimed that evolution, properly understood, justified a laissez-faire social ideology, often now inappropriately attributed to Darwin (Spencer 1851, 1852a, 1852b, 1864). His claims that nature exhibited inherent values that should guide human society were sharply criticized by philosopher G.E. Moore (1903), who famously called Spencer's error the naturalistic fallacy. What is the difference between facts and values? Why do facts alone not yield or justify values? How is understanding or explaining a behavior not the same as endorsing it?
How do even universal facts about humans (or "human nature") not dictate human ideals, values or notions of what should be? In what ways is frequency of occurence (or normality) not the same as a justified value? How do "is" and "ought" (description and prescription) differ?
Based on this analysis, why is it not possible for evolutionary theory or science to be a source of "natural" ethics or moral principles?
Target Concept: Neither nature nor evolution expresses or justifies values or particular ethical principles. |

[ 55 ] |
Spencer's claims seemed misguided on an even more fundamental level. His psychology and sociology reflected a biological determinism and progressive ideology based ultimately on his own political beliefs. He did not extract values from nature, so much as inscribe them into his own scientific theories. This is called the naturalizing error. It is an error in science. Scientists may do this without realizing it, with their cultural perspective functioning like a conceptual blindspot. Here is a 1908 painting by Henri Rousseau. Rousseau sought realism and intentionally based his depictions on greenhouse specimens. But they are highly idealized and romanticized, nonetheless. How do we guard against a "scientific" account of nature becoming tainted with a cultural artifact? In what ways might viewing nature as fundamentally competitive and ruthless— or even as morally perfect — be shaped by our economic and cultural views more than by critical interpretation of the evidence?
Target Concept: Cultural bias may generate error in science, with adverse effects beyond science. |
 
[ 56 ] |
If humans have largely open behavioral programs, what does this imply about human choice and potential? Is human behavior fully determined (or predictable)? Profile a plausible biological perspective on why philosophers have disagreed for centuries, and continue to disagree, about the basics of ethics.
Target Concept: Biologically, humans have multiple moral potentials. |
|
References
- Allchin, D. 1992. Simulation and analysis of information-center foraging. Behaviour 122, 288-305.
- Allchin, D. 2008. Naturalizing as an error-type in biology. In Proceedings of the Sixth Brazilian Conference for the History and Philosophy of Biology.
- Axelrod, R. 1984. The Evolution of Cooperation. New York, NY: Basic Books.
- Baglione, V., Canestrari, D., Marcos, J.M., Ekman, J. 2003. Kin selection in cooperative alliances of carrion crows. Science 300: 1947-1949.
- Barrett, P., Gautrey, P.J., Herbert, S., Kohn D. & Smith, S. 1987. Charles Darwin's Notebooks (1836-1844). Ithaca, NY: Cornell University Press.
- Bils, J. and Singer, S. 1996. Gorilla saves tot in Brookfield Zoo ape pit. Chicago Tribune (Aug 17.), 11.
- Boehm, C. 1999. Hierarchy in the Forest: The Evolution of Egalitarian Behavior. Cambridge, MA: Harvard University Press.
- Bradie, M. 1994. The Secret Chain. Albany, NY: State University of New York Press.
- Bshary, R. and Gutter, A.S. 2006. Image scoring and cooperation in a cleaner fish mutualism. Nature 441, 975-978.
- Burkart, J.M., Fehr, E., Efferson, C. and van Schaik, C.P. 2007. Other-regarding preferences in a non-human primate: common marmosets provision food altruistically. Proc. Nat. Acad. Sci., U.S. 104, 19762-19766.
- Camazine, S., Deneubourg, J.-L., Franks, N. R., Sneyd, J., Theraulaz, G., & Bonabeau, E. 2001. Self-Organization in Biological Systems. Princeton, NJ: Princeton University Press.
- Campbell, D.T. 1974. Evolutionary epistemology. In P.A. Schilpp (ed.), The Philosophy of Karl Popper. La Salle, IL: Open Court, pp. 413-63.
- Clutton-Brock, T.H. & Parker, G.A. 1995. Punishment in animal societies. Nature, 373, 209-216.
- Clutton-Brock, T. 2002. Breeding together: kin selection and mutualism in cooperative vertebrates. Science 296, 69-72.
- Clutton-Brock, T.H., O'Riain, M.J.O., Brotherton, P.N.M., Gaynor, D., Kansky, R., Griffin, A.S. and Manser, M. 1999. Selfish sentinels in cooperative mammals. Science 284, 1640-1644.
- Dally, J. et al. 2006. Food-caching western scrub-jays keep track of who was watching when. Science 312, 1662-16XX.
- Damasio, H., Grabowski, T., Frank, R., Galaburda, A.M. and Damasio, A.R. 1994. The return of Phineas Gage: clues about the brain from the skull of a famous patient. Science 264, 1102-1105.
- Darwin, C. 1871. The Descent of Man. London: John Murray.
- Darwin, C. 1872/1965. The Expression of the Emotions in Man and Animals. Chicago, IL: University of Chicago Press.
- Emlen, S.T., 1978. The evolution of cooperative breeding in birds. In J.R. Krebs and N.B. Davies, eds., Behavioural Ecology: An Evolutionary Approach, Oxford, UK: Basil Blackwell, pp. 245-281.
- Farber, P. 1994. The Temptations of Evolutionary Ethics. Berkeley, CA: University of California Press.
- Fehr, E. and Gächter, S. 2002. Altruistic punishment in humans. Nature 415, 137-140.
- "Gorilla at an Illinois Zoo Rescues a 3-year Old Boy." New York Times (August 17, 1996).
- Gould, S.J. 1981. The Mismeasure of Man. New York, NY: W.W. Norton.
- Gould, S.J. 1983. The guano ring. In Hen's Teeth and Horse's Toes, New York, NY: W.W. Norton, pp. 46-55.
- Grafen. 1990. Do animals really recognize kin? Animal Behaviour 39, 42-54.
- Greene, J.D., Sommerville, R.B., Nystrom, L.E., Darley, J.M. and Cohen, J.D. 2001. An fMRI investigation of emotional engagement in moral judgment. Science 293:2105-2108.
- Greene, J.D. and Haidt, J. 2002. How (and where) does moral judgment work? Trends in Cognition 6, 517-523.
- Güreck, Ö., Irlenbusch, B. and Rockenbach, B. 2006. The competitive advantage of sanctioning institutions. Science 312, 108-111.
- Hamilton, W.D. 1964. The genetical evolution of social behavior. J. Theoretical Biology 7, 1-52.
- Hardin, G. 1968. The tragedy of the commons. Science 162, 1243-1248.
- Harlow, J.M. 1868. Recovery from the passage of an iron bar through the head. Publ. of the Massachussetts Medical Society 2, 328-347. Facsimile reprinted in Macmillan (2000), pp. 401-422.
- Hauert, C., Traulsen, A., Brandt, H., Nowak, M.A. and Sigmund, K. 2007. Via freedom to coercion: the emergence of costly punishment. Science 316, 1905-1907.
- Hauser, M.D. 1992. Costs of deception: cheaters are punished in rhesus monkeys. Proc. Nat. Acad. Sci., U.S. 89, 12137-12139.
- Heinrich, J., McElrath, R., Barr, A., Ensminger, J., Barrett, C., Bolyanatz, A., et al. 2006. Costly punishment across human societies. Science 312: 1767-1770.
- Hermann, B., Thöni, C. and Gächter, S. 2008. Antisocial punishment across societies. Science 319, 1362-1367.
- Hofstadter, D.R. 1979. Golden, Escher, Bach. New York, NY: Basic Books.
- Holden, C. 2008. Smart birds lend a beak for food. Science 319, 1746.
- Holland, J. H. 1998. Emergence: From Chaos to Order. Cambridge, MA: Perseus.
- Jarvis, J.U.M. 1981. Eusociality in a mammal: cooperative breeding in naked mole-rat colonies. Science 212, 571-573.
- Krebs, J.R. and Davies, N.B. 1993. An Introduction to Behavioural Ecology. Oxford, UK: Blackwell Scientific.
- Langford, D.J., Crager, S.E., Shehzad, Z., et al. 2006. Social modulation of pain as evidence for empathy in mice. Science 312:1967-1970.
- Lewontin, R.C. 1993. Biology as Ideology: The Doctrine of DNA. New York, NY: Harper Collins.
- Lewontin, R.C., Rose, S. & Kamin, L.J. 1984. Not in Our Genes. New York, NY: Pantheon Books.
- Lieberman, D., Tooby, J. and Cosmides, L. 2003. Does morality have a biological basis? An empirical test of the factors governing moral sentiments relating to incest. Proc. Roy. Soc. 270, 819-826.
- Lieberman, D., Tooby, J. and Cosmides, L. 2007. The architecture of human kin detection. Nature 445, 727-731.
- Macmillan, M. 2000. An Odd Kind of Fame: Stories of Phineas Gage. Cambridge, MA: MIT Press.
- Masserman, J.H., Wechkin, S. and Terris, W. 1963. 'Atruistic' behavior in rhesus monkeys (Macaca mulatta). Proc. Nat. Acad. Sci., U.S. 89:12137-12139.
- Melis, A.P., Harem B., and Tomasello, M. 2006. Chimpanzees recruit the best collaborators. 311:1297-1300.
- Milinski, M. and Rockenbach, B. 2007. Spying on others evolves. Science 317, 464-465.
- Miller, G. 2005. Reflecting on another's mind. Science 308, 945-947.
- Miller, G. 2008. The roots of morality. Science 320, 734-737.
- Moore, G.E. 1903. Principia Ethica. Cambridge, UK: Cambridge University Press.
- Murphy, N. and Brown, W.S. 2007. Did My Neurons Make Me Do It?: Philosophical and Neurobiological Perspectives on Moral Responsibility and Free Will. Oxford, UK: Oxford University Press.
- Niendenthal, P.M. 2007. Embodying emotion. Science 316, 1002-1005.
- Nowak, M.A. 2006. Five rules for the evolution of cooperation. Science 314:1560-1563.
- Nowak, M.A. and Sigmund, K. 1998. Evolution of indirect reciprocity by image scoring. Nature 93, 573-.577.
- Nowak, M.A. and Sigmund, K. 2005. Evolution of indirect reciprocity. Nature 437, 1291-1298.
- O'Riain, M.J. and Faulkes, C.G. 2008. African mole-rats: eusociality, relatedness and ecological constraints. In Ecology of Social Evolution, ed. Judith Korb and Jörgen Heinze (Berlin: Springer), 207-223.
- Pennisi, E. 2006. Social animals prove their smarts. Science 312, 1734-1738.
- Pfaff, D. 2007. The Neuroscience of Fair Play. New York NY: Dana Press.
- Ratiu, P. and Talos, I.-F. 2004. The tale of Phineas Gage, digitally remastered. New England Journal of Medicine 351(23, Dec. 2): e21.
- Richerson, P.J. and Boyd, R. 2005. Not By Genes Alone: How Culture Transformed Human Evolution. Chicago, IL: University of Chicago Press.
- Rose, S. 1997. Lifelines: Life Beyond the Gene. Oxford, UK: Oxford University Press.
- Seed, A., Clayton, N. and Emery, N. 2008. Proc. Roy. Soc.
- Shelley, E.L. and Blumstein, D.T. 2004. The evolution of vocal alarm communication in rodents. Behavioral Ecology 16, 169-177.
- Sherman, P.W. 1977. Nepotism and evolution of alarm calls. Science 197, 1246–1253.
- Sherman, P.W., Jarvis, J.U.M., Alexander, R.D. 1991 The Biology of the Naked Mole-Rat. Princeton, NJ: Princeton University Press.
- Sinervo, B. and Clobert, J. 2003. Morphs, dispersal behavior, genetic similarity, and the evolution of cooperation. Science 300, 1949-1951.
- Sober, E. & Wilson, D.S. (1998). Unto Others. Cambridge, MA: Harvard University Press.
- Spencer, H. 1851/1969. Social Statics. London: John Chapman. Reprinted by Augustus M. Kelley, New York, NY.
- Spencer, H. 1852a. A theory of population, deduced from the general law of animal fertility. Westminster Review 57, 468-501.
- Spencer, H. 1852b. The development hypothesis. The Leader (March 20).
- Spencer, H. 1864/1924. First Principles, 6th ed.., New York, NY: Appleton and Company.
- Sterelny, K. 2001. The Evolution of Agency and Other Essays. Cambridge, UK: Cambridge University Press.
- Trivers, R.L. 1971. The evolution of reciprocal altruism. Quarterly Review of Biology 46, 35-57.
- de Waal, F. 1989. Food sharing and reciprocal obligations among chimpanzees. J. Human Evol. 18, 433-459.
- de Waal, F. 1996a. Good Natured: The Origins of Right and Wrong in Humans and Other Animals. Cambridge, MA: Harvard University Press.
- de Waal, F. 1996b. op-ed. New York Times (August 23).
- de Waal, F. 2005. Our Inner Ape. New York, NY: Riverhead Books.
- de Waal, F. and Aureli, F. 1996. Reconciliation, consolation and possible cognitive differences between macaques and chimpanzees. In Reaching into Thought, ed. A.E. Russon and S.T. Parker (Cambridge, UK: Cambridge University Press), 80-110.
- de Waal, F. Leimgruber,K. and Greenberg, A.R. 2008. Giving is self-rewarding for monkeys. Proc. Nat. Acad. Sci. U.S. 105, 13685–13689.
- Warneken, F. and Tomasello, M. 2006. Altruistic helping in human infants and young chimpanzees. Science 311, 1301-1303.
- Wilkinson, G.S. 1984. Reciprocal food sharing in the vampire bat. Nature 308, 181-184.
- Wilkinson, G.S. 1988. Reciprocal altruism in bats and other mammals. Ethology and Sociobiology 9, 85-100.
- Wilkinson, G.S. 1990. Food sharing in vampire bats. Scientific American (Feb.): 76-82.
- Woolfenden, G.E. and Fitzpatrick, J.W. 1978. The inheritance of territory in group breeding birds. BioScience 28, 104-108.
- Woolfenden, G.E. and Fitzpatrick, J.W. 1984. The Florida Scrub Jay: Demography of a Cooperative- breeding Bird. Princeton, NJ: Princeton University Press.
- Wright, J., Maklakov, A.A., Khazin, V. 2001. State-dependent sentinels: an experimental study in the Arabian babbler. Proc. Roy. Soc. B, 268, 821-826.
- Zimmer, C. 2003. How the mind reads other minds. Science 300, 1079-1080.
|
|
